徕卡共聚焦课堂第5讲:荧光和量子点的基本原理和发展历史
在您的科研生涯的某个时候,都有可能会用到荧光显微镜。这种无处不在的技术改变了显微镜学家对研究对象进行成像、标记和追踪的方式,不论是整个生物体,还是单个蛋白质等等。
通过本文,我们将探讨什么是“荧光”,包括其定义背后的历史和基础物理原理,绿色荧光蛋白(GFP)的发现和应用,并展望量子点等荧光探针不断扩大的应用领域。
我们如何定义“荧光”?
在任何搜索引擎中输入“荧光的定义”,你会得到以下语句,或者非常相似的内容;
“由较短波长的入射辐射,如X射线或紫外线,某些物质会发出可见或不可见的辐射。”
这个定义与荧光显微镜(图1)有何关联呢?“波长较短的入射辐射”只是用来“激发”样品发射荧光的光源。这种光线包括可见光、紫外线(UV)和红外线(IR),显微镜的光源可以是汞或氙弧光灯,也可以是激光。荧光基团(即上述定义中的“某些物质”)是具有特殊性质的化合物,它们在较短波长光线的激发下,可以再次发射较高波长的光线/光子。

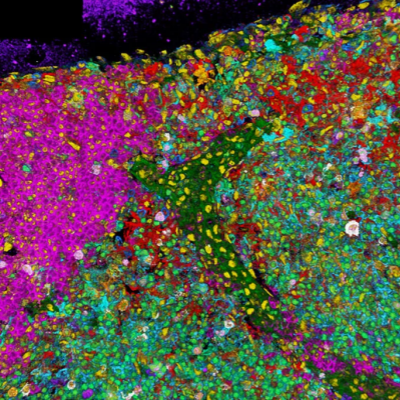
图1:荧光显微镜下显示的荧光标记细胞。
波长的基本单位是米,“波长”定义为光波的两个连续波峰或波谷之间的距离。波长的符号是希腊字母λ(l)。显微镜使用的波长通常以纳米(nm)为单位,可见光谱段在400至700纳米之间,紫外光谱段在400纳米以下,红外光谱段从700纳米开始(图2)。

图2:可见光光谱
荧光基团按其激发和发射波长分类,通常以图形的形式显示最大波峰。荧光基团在所谓的“基态”中自然稳定。它们吸收到(来自“较短波长”入射光的)光子时,光子的能量将荧光基团的电子提升到更高能量的“激发”状态。被激发的电子不会保持在这种状态,而会失去振动能量,并在返回基态时发射出一个较长波长的光子。对于显微镜使用荧光基团,从激发到返回基态的一个完整周期需要0.5到20纳秒。
荧光基团的激发和发射波长通常缩写为希腊字母lambda,加上下标“ex”(激发)或“em”(发射;即lex或lem)。
每个荧光基团都有一个不同的最大激发和发射谱段,这一特性用来区分同一样品中的不同标靶。例如,使用荧光染色剂DAPI(4',6-二脒基-2-苯基吲哚)高亮显示细胞中的核蛋白,同时使用荧光标记的鬼笔环肽来高亮显示肌动蛋白细胞骨架。
荧光的雅布隆斯基图
波兰物理学家亚历山大·雅布隆斯基(Aleksander Jablonski,1898-1980)首先用三能级能量图描述了基态/激发/发射之间的循环,因此称为“雅布隆斯基图”。 1930年,他凭借题为《关于激发光波长变化对荧光光谱的影响》的论文获得华沙大学博士学位。1933年,他在《自然》杂志上发表了自己的论文(《染料中反斯托克斯荧光的效率》”),其中含有雅布隆斯基图。这种简单而有效的示意图清晰表现出荧光基团从基态到激发态的激发行为,然后在发射较长波长光子后回到基态(图3)。
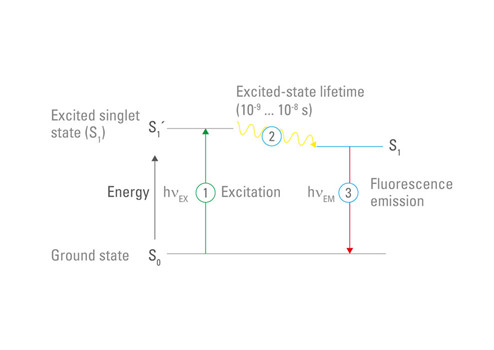
图3:荧光雅布隆斯基图。荧光基团会吸收光子。光子的能量将荧光基团的电子提升到更高能量的“激发”状态。随后,激发的电子失去振动能量,并在返回基态时发射一个较长波长的光子。
虽然图3是简化版的雅布隆斯基图,但应该可以注意到,荧光基团可以存在多个不同的激发和发射状态。此外,荧光基团可以通过不同的弛豫状态返回基态,这些弛豫状态被称为“三重态”,具体取决于分子的电子自旋。
斯托克斯位移
乔治·加布里埃尔·斯托克斯爵士(1819-1903)是爱尔兰数学家和物理学家,他一生中在光学和光线领域颇有建树。1849年他受命担任剑桥彭布罗克学院的卢卡斯数学教授,并一直担任该职务,直到1903年去世。他写了一篇文笔优美的描述性论文,发表于1852年,名为《论光的折射性的变化》[1]。他在论文中使用的语言不同于我们在现代科学文献中使用的语言。以下是斯托克斯爵士所用精彩语言的两段简短摘录;
“去年深秋,虽然因为节气迟来,观察机会不多,我从不同渠道得知,之前提到过的那种具有高内色散性的黄色玻璃采用氧化铀染色。”
以及:
“看到试管在浸入不可见光线的瞬间点亮,这当然是一个奇怪的景象:那实际上是可见的黑暗。总的来说,这种现象有点不可思议。”
“斯托克斯位移”这词就是为了纪念这位科学家。当激发的电子回到基态并发出光子时,其波长始终大于激发荧光基团的入射光线的波长。这是由于光的特性,即波长与辐射能量成反比。斯托克斯位移描述了荧光基团的峰值激发波长和峰值发射波长之间的差值(以纳米为单位)(图4)。
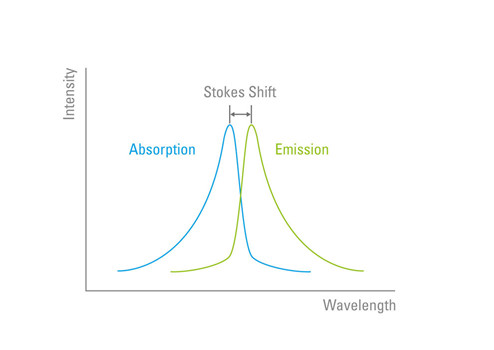
图4:斯托克斯位移发射光的波长始终大于用来激发荧光基团的光线波长。
增加斯托克斯位移,荧光基团的激发光和发射光之间区别就越明显。荧光基团的电子排布和分子结构令其在斯托克斯位移方面有着独特的性质。
绿色荧光蛋白的发现与应用
斯托克斯爵士首先使用“荧光”这个术语来描述他观察到的现象,但其发现可以追溯到1565年。来自西班牙的医生兼植物学家尼古拉斯·莫纳德斯(Nicolas Monardes)描述了一种墨西哥树(Lignum nephriticum),其木头被注入了奇怪的蓝色。尽管有这些早期的观察,但直到约400年后,人们才在活体组织中找到一种绿色荧光物质。1955年,Davenport和Nicol发表论文[2],描述了一种在称为水螅水母(Hydromedusae)的水母亚纲生物的嗜酸性粒细胞中找到的发光组织。当时,两位论文作者并没有意识到这些嗜酸性粒细胞含有绿色荧光蛋白(GFP)。
直到1962年,下村脩(Osamu Shimomura,出生于1928)发表了一篇论文[3],认定这种发光成分是一种蛋白质。通过与他在普林斯顿大学的老师(弗兰克·约翰逊教授)合作,他们从生物发光水母Aequorea Victoria(维多利亚多管发光水母)身上收集到样本。他们用一种独特的方法从水母中分离出荧光蛋白,即通过棉布袋挤压分离的生物发光组织,产生一种被下村称为“挤压物”的溶液。
1994年,哥伦比亚大学的马丁·查尔菲(Martin Chalfie,出生于1947年)教授发表论文,证明了基因编码GFP可以在原核细胞和真核细胞(即大肠杆菌和秀丽隐杆线虫的神经元)中实现功能表达[4]。该论文写道“由于这种荧光不需要外源底物和辅助因子,GFP表达可以用于监测生物体内的基因表达和蛋白质定位。”正是这篇文章的发表为GFP在生物学研究中的广泛应用铺平了道路。
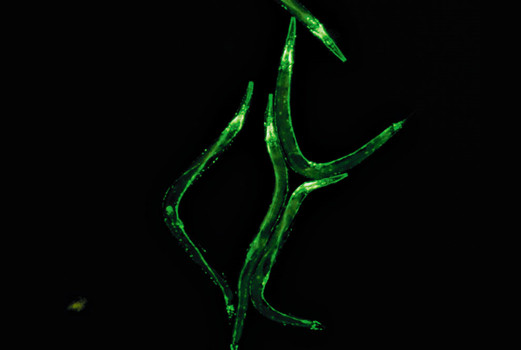
图5:线虫,GFP在神经系统中的表达。
一年后,一直在研究野生型GFP突变体的加州大学圣迭戈分校教授钱永健(1952-2016)在《自然》的科学通讯上发表了自己的论文[5]。钱教授和他的同事发现,他们选择的一个单点突变(S65T)具有最长的激发和发射波长(490/510nm),这令它更具有光稳定性,并带来众所周知的GFP激发/发射峰。
2008年,下村脩、钱永健和查尔菲因在GFP的发现和应用方面发挥的重要作用共同获得诺贝尔化学奖。下村脩在他的诺贝尔演讲中表示,“当我在1979年发现GFP的发色团时,我认为自己已经完成了所有研究工作,因而决定终止我在GFP方面的工作,以便集中精力研究生物发光”。他接着承认,“GFP是一种美丽的蛋白质,但在发现之后的30年中,它依然百无一用。”
自从钱永健发现GFP的应用之后,他的实验室已经设计出多种GFP变体,涵盖了大部分的可见光谱,并彻底改变了光学显微镜和成像领域。得益于这种基因工程,GFP的色彩范围从光谱的蓝色谱段(EBFP;380/460纳米)到光谱的黄色谱段(YFP;514/527纳米)。
报告基因和探针
除了成像领域,荧光团可以作为研究细胞和生物体中基因表达的“报告基因”。荧光素酶以及基因编码GFP,就是检查细胞是否表达特定基因的常用报告基因。生物体或细胞被基因改造的病毒DNA或质粒转染,当目标基因获得表达时,这些质粒会发出荧光。细胞内部的相关细胞器或位点可以在受转染的细胞中进行观察和检查。
利用荧光基团标记(或“标注”)的抗体通常称为“荧光探针”。在GFP和其他荧光基团得到广泛应用之前,两种最常用的荧光基团标记抗体是FITC(异硫氰酸荧光素)和TRITC(异硫氰酸四甲基罗丹明)。尽管FITC和TRITC仍在使用,但该领域已取得重大进展,至少可以说,荧光基团和探针的选择非常广泛。
当然,如今实验室中使用最为广泛的荧光基团之一是“Alexa Fluor”染色剂。目前Alexa Fluor染色剂的可用色彩范围已超越可见光谱,其中激发波长范围可达346至784纳米。
量子点
1981年,俄罗斯科学家阿列克谢·埃基莫夫(Alexey Ekimov)在圣彼得堡瓦维洛夫国立光学研究所工作时,首次在玻璃基质中发现了量子点。然而,在新泽西州AT&T贝尔实验室从事半导体工作的路易斯·布鲁斯(Louis Brus)首先发现了量子点胶体溶液。他现在是哥伦比亚大学化学系教授。在1983年和1984年发表的两篇论文中,布鲁斯将量子点描述为“小型半导体微晶”。
量子点有着奇特的行为方式,尽管它们包含100到10万个原子,但它们表现出的性质就像它们由单个原子组成一样。当然,它们确实符合荧光基团的性质,即它们可以吸收光能并激发,然后在返回基态时释放光子。不过,量子点发射光线的波长取决于量子点的大小,即量子点越小,发射波长就越短。这种特性是因为较小的量子点具有较大的“最小带隙”。这正是激发电子到更高能态所需的能量。由于较小的量子点需要更多的激发能量,它们随后会发射较短波长的光(波长与激发能量成反比)。
大约20年后,即2002年,加州量子点公司的生命科学研究人员实现了量子点的商用开发。
量子点是一种非常明亮且稳定的荧光工具。研究表明,量子点的的亮度比传统荧光基团高几个数量级,尽管与有机染料相比,量子点的明亮程度存在一些差异[6]。就光稳定性而言,量子点的稳定性是传统荧光基团的100倍,在一项活体成像研究中,量子点的荧光持续时间长达4个月[7]。
在传统荧光探针中,一个或多个荧光基团可以附着在单个相关抗体上。然而,由于量子点的表面积非常大,许多分子和抗体可以附着在单个量子点上,这会带来许多优点,包括荧光信号放大。量子点的使用也为多重分析提供了优势,可以在检测中同时对各种波长进行成像。在这种分析中,唯一的变量是研究者选择的量子点大小(因此发射波长)。可以使用白光同时激发大范围的量子点,这样就可以避免使用多道激光和多次调整来形成最终影像。